
Open Access, Volume 9
Brain functions of saffron and crocin: Case report and literature review
Shiho Kito1; Yukihiro Shoyama2*
1Association for Health Economics Research and Social Insurance and Welfare, Tokyo, Japan.
2Faculty of Pharmacy, Nagasaki International University, Sasebo, Nagasaki, Japan.
Yukihiro Shoyama
Faculty of Pharmacy, Nagasaki International University, Sasebo, Nagasaki, Japan.
Email: shoyama@niu.ac.jp
Received : July 31, 2023,
Accepted : Sep 20, 2023
Published : Sep 22, 2023,
Archived : www.jclinmedcasereports.com
Abstract
Saffron is known to contain approximately 150 components, among which crocetin glycosides, i.e. crocin, safranal and picrocrocin have been found to be the active substances. In recent years, research on the anticognition activity of saffron has been widely conducted, and it has become clear that the active ingredient is crocin. Crocin is a component of crocetin with four glucose bonds, but as crocetin is a diterpene linked by conjugated double bonds, it is sensitive to oxygen, heat and light, causing secondary changes and degradation. For this reason, quality control is essential for researches. In-vitro experiments have shown that crocin has antioxidant and anti-inflammatory-lowering effects, lowered ceramide levels, increased glutathione levels, lowered amyloid-β, increased acetylcholine and dopamine levels and increased long-term potentiation. In in-vivo experiments, crocin increased memory learning capacity and activated long-term potentiation. Crocin also induced sleep effects, particularly non-REM sleep. Saffron was also found to have sleep effects in clinical studies. Epidemiological studies revealed that these effects were linked to anti-cognitive effects, so saffron and crocin can be said to be indirectly effective against Alzheimer’s disease. The clinical trial of saffron extract ameliorated symptoms in Alzheimer’s patients. From these findings saffron and crocin can potentially be applied for treatment and prevention of dementia in patients.
Keywords: Saffron; Crocin; Quality control; Mechanism; Anti-dementia activity.
Copy right Statement: Content published in the journal follows Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0). © Shoyama Y (2023)
Journal: Open Journal of Clinical and Medical Case Reports is an international, open access, peer reviewed Journal mainly focused exclusively on the medical and clinical case reports.
Citation: Kito S, Shoyama Y. Brain functions of saffron and crocin: Case report and literature review. Open J Clin Med Case Rep. 2023; 2119.
Introduction
Materia Medica described by Dioscorides indicates that saffron is the fresher the better, and is used for poor circulation and uterine medicine, for bowel movements and as a tonic. In Greek time it was used as an aphrodisiac. Saffron was brought to China via India around the 13th century and has been used medicinally until today. ‘Honzo-gangmoku’ published by Li Shi-zhen indicated that saffron is used to treat a depressed and anguished mind that cannot be dissipated by blood. It has been used since ancient times for depression, breathing problems, haematemesis, chills, hysteria, fear, ecstasy, female menopause, postpartum bleeding and abdominal pain.
Saffron belongs to the iris family and is a perennial herb that is sterile and therefore propagates from bulbs. The three pistils of each flower are collected and dried to prepare saffron. Five kg of fresh pistils (Figure 1) are harvested from 90,000-100,000 saffron flowers resulting in 1 kg of dried saffron. This labour-process makes it the most expensive herb medicine.

Figure 1: Flowering of saffron (yellow arows indicate pistil).
Saffron has been used since ancient times as a medicine, spice and dye. Approximately 150 constituents have been determined in saffron, such as terpenoids, amino acids, anthraquinones, alkaloids and others. Among them crocin, safranal, picrocrocin and crocetin glycosides are major constituents [1]. Carotenoid pigments include crocetin with four molecules of glucose, crocin, three molecules of glucose and two molecules of glucose. Flowering state and the crocetin glycoside relationship were investigated, and the results show that the crocetin glycoside content increases as the flowering stage and reaching a maximum at full bloom, and decreased by nearly 2% after full bloom [2]. The quality is also known to fluctuate depending on the weather conditions at the time of collection.
Quality control of saffron and its constituent, crocin
Dioscorides showed that saffron is the fresher the better. This could mean that saffron undergoes compositional changes during storage, resulting in a loss of potency. The principal crocetin glycosides of saffron are crocin, crocetin gentiobiose glucose ester (crocin 2) and crocetin di-glucose ester (crocin 3), with respective contents of about 14,5 and 0.7%, respectively but during storage crocin 3 apparently increase. This indicates that the endogenous β-glycosidase is active and cleaves the glycosidic bonds in the presence of moisture resulting in crocin 2 to crocin 3. It was found that the double bonds of the crocetin molecule were cleaved by various storage conditions, i.e. higher temperature, oxygen, etc., resulting in the degradation of crocin to unknown compounds [2] as indicated in Figure 2.
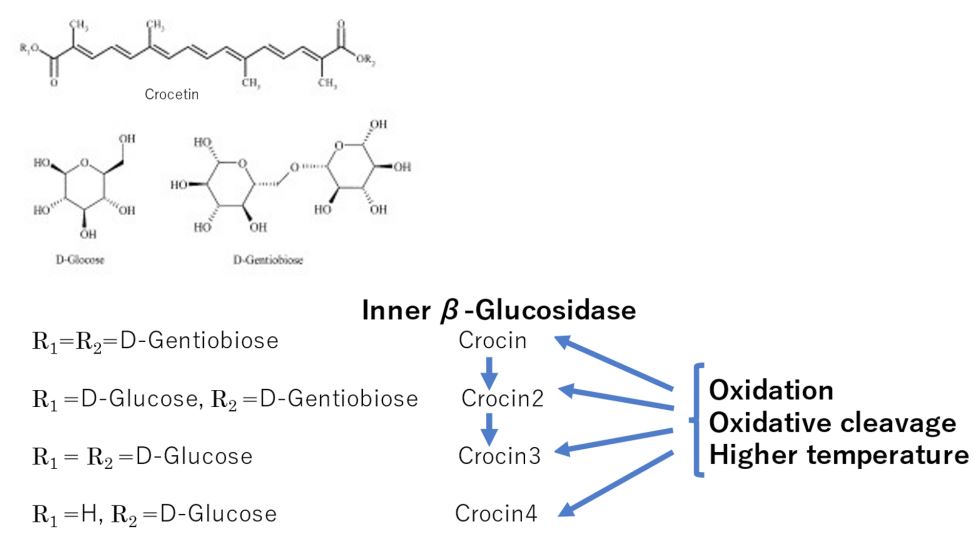
Figure 2: Major constituents in saffron and artificial degradation of crocetin glycosides.
The results show that the content of crocin is fluctuates considerably depending on the storage conditions. Therefore, at least storage at -20°C under dry conditions is necessary to prevent such degradation [2-4]. More recently Alehossseini et al. investigated the stability of saffron extract through encapsulation in zein protein [5]. When conducting pharmacological researches with saffron, the quality control with a focus on crocin is essential since it has been proven that the active component in saffron with regard to brain function is crocin [6]. Moreover, the strength of long-term potentiation (LTP) is strongest for crocin with 4 glucose, and weakens as the number of glucose decreases in the order crocin 3 and crocin 2 (see Figure 2) [7]. This tendency has been observed in pharmacological activity studies of naturally occurring glycosides such as cardiac steroids [8], ginseng saponin [9], saikosaponin [10] and haemolytic saponin [11]. For this reason, the quality control of crocin in saffron is necessary for researches using saffron. With the aim of developing a simple and rapid analytical method, a monoclonal antibody (MAb) against the crocin was developed [12].
Mechanism of anti-cognitive activity for crocin
In order to confirm the anti-oxidant property of crocin the effect of crocin on PC-12 cells under serum and glucose-free conditions was compared with that of α-tocopherol which is well-known anti-oxidant compound. Phosphatidylserine (PS), normally present in the endothelium of cells, is translocated to the outer membrane of cells by oxidative stress, and this translocation is considered an early signal for apoptosis induction. FITC-bound AnnexinV binds to negatively charged PS and exhibits a ring-shaped fluorescence. PC-12 cells under serum and glucose-free conditions clearly show ring-shaped fluorescence. No ring fluorescence is observed in cells in medium with 10 μM of crocin. On the other hand 10 μM of α-tocopherol detects rings, although the number is small. It is clear that the cells in the crocin-added medium maintained a more normal state when compared to those in the α-tocopherol added medium [13,14]. Oxidative stress is known to cause many brain disorders, and is deeply involved in Alzheimer's disease, Parkinson's disease, and Hutchinson's disease as diseases of the Central Nervous System (CNS). Since the antioxidant effect stronger and safer than that of α-tocopherol, it is considered to be applicable to neurological disorders, and further animal and clinical experiments are desirable. Recent studies have shown that apoptosis is induced by oxygen deficiency in the brain and brain damage, and that apoptosis also occurs in the brain of Alzheimer's patients. Recently Wang et al. certified that the oxidative stress is associated with apoptosis signaling pathway [15].
With regard to apoptosis of PC-12 cells, it is known that removal of serum and Nerve Growth Factor (NGF) causes an increase in ceramide levels inducing apoptosis. PC-12 cells remain normal in medium containing serum and glucose, but when serum and glucose are removed from the same medium, 60% of cell death were confirmed by the Trypan blue dye method. In contrast, the addition of 10 μM crocin significantly inhibited the morphological phenomenon showed that crocin blocked TNF-α-induced PC-12 cell death resulting in 85% survival compared to control cells [16].
Serum deprivation increased intracellular ceramide levels in HN9.10e cells resulting apoptosis. This finding suggests that a similar phenomenon in PC-12 cells will be occurred. In fact the ceramide concentration significantly increased like as 3.5-fold by culturing in serum and glucose free medium compared to the control medium [17].
Sphingomyelinase acts on sphingomyelin to produce ceramide, and sphingomyelinase activity in PC-12 cell homogenates was measured, reaching a maximum at 1 hr and decreasing to control levels at 3 hr. Next, sphingomyelinase activity increased markedly under serum and glucose-free conditions, but the addition of crocin decreased the activity in a dose-dependent manner. However, no direct effect of crocin on sphingomyelinase activity was observed [13]. The SAPK/JNK signaling system and the sphingomyelin pathway might be conjugated to give a mixed function by stress in U937 and BAE cells resulting that environmental stress conditions activated the stress-activated protein kinase SAPK/JNK cascade in PC-12 [18]. Rukenstein et al. found that serum and glucose deprivation medium activated JNK phosphorylation in PC12 cells nearly 4-fold when compared with the control cells [19].
Glutathione (GSH) levels in PC-12 cells under serum and glucose-free conditions were measured. GSH levels were also measured under crocin and crocin-free conditions. 3 hr of treatment of PC-12 cells under serum and glucose-free conditions resulted in half the concentration of GSH compared to the control. On the other hand, the addition of crocin increased GSH levels in a dose-dependent manner, and GSH levels remained higher than those of controls after 3 hr. Furthermore, the effect of crocin on glutathione synthetase was investigated resulted that crocin increased GSH which decreased oxidation stress resulting the prevention of neuronal cell apoptosis and learning and memory obstacle by activating the enzymes [20]. Next Sugiura et al. evaluated the activity of crocin in the dentate gyrus of rat hippocampal slice using ethanol as a LTP inhibitor. LTP is significantly suppressed by 50-75 nM alcohol administration. This suppression of LTP by alcohol was found to be dose-dependently enhanced by the pre-administration of crocin 10-30 μM 5 min before alcohol administration although crocin alone did not attenuate the LTP [21]. Table 1: Summarized the mechanism of neuronal function of crocin. In addition to the above, crocin has been reported to exhibit brain functions as shown in Table 1.
Table 1: Mechanism of action of crocin on brain function in-vitro.
| Neuronal cell apoptosis | Reference |
|---|---|
| Oxidation | 13-15 |
| Inflammation | 22,23 |
| Ceramide level | 16-19 |
| Glutathione level | 20 |
| Learning and memory | |
| Acetylcholine | 24 |
| Amyloid B-peptide | 25,26 |
| Dopamine | 27,28 |
| Long-term potentiation | 21 |
Effects of saffron and crocin on alcohol-impaired memory learning and long-term potentiation invivo
Although ethanol, acetaldehyde, scopolamine, hyoscine or amyloid β (Aβ) and so on is used as an inhibitor in memory learning and LTP experiments, ethanol is generally used because of its high sensitivity and convenient weighing. The Saffron Extract (SE) activity on memory in aged mice was associated with the antioxidant activity [29]. Roustazade et al. found that the SE dose reflects the activity of brain function in stressed rats [30]. The SE were studied on learning and memory in Step Through (ST) and Step Down (SD) tests in normal mice as well as in learning and memory-impaired mice. A single administration of SE had no effects on memory registration, consolidation or retrieval in normal mice. SE (125-500 mg/ kg) dose-dependently reduced the ethanol-induced impairment of memory registration both in ST and SD tests and the 30% of ethanol-induced impairment of memory retrieval in SD test. The SE decreased the motor activity and prolonged the sleeping time induced by hexobarbital resulting that SE ameliorate the impairment effects of ethanol on learning and memory processes, and possesses a sedative effect [31]. Since SE showed ameliorative effects on alcohol-blocked memory learning deficits, Morimoto et al. purified by silica-gel column chromatography using activity-guided separation to reach crocin as active principle. This result is easily supposed, given that crocin is a major constituent, around 40% of the SE [2,7].
It became evident that no effect on memory possession by a single-oral investigation of crocin in normal mice was found in ST and SD tests, respectively. If crocin was administered 10 minutes before the ethanol injection, the latency in the ST test dose-dependently increased compared with that of ethanolinjected mice. The investigation of 200 mg/kg crocin increased the number of successful mice decreasing the error numbers compared to the ethanol-treated group in the test trial [32].
Sugiura et al. evaluated the activity of crocin in the dentate gyrus of rat hippocampal slice using ethanol as a LTP inhibitor. LTP is significantly suppressed by 50-75 nM alcohol administration. This suppression of LTP by alcohol was found to be dose-dependently enhanced by the pre-administration of crocin 10-30 μM 5 min before alcohol administration although crocin alone did not attenuate the LTP [21].
Since LTP is an important phenomenon related to short-term memory, Sugiura et al. evaluated the activity of SEs in the rat hippocampal using ethanol as a LTP inhibitor. LTP does not occur when SE alone is administered orally. On the other hand, the administration of SE prior to ethanol administration clearly reduced the inhibition of LTP expression by ethanol. SE at 125-250 mg/kg was found to exhibit dosedependent manner [33].
Effect of saffron and crocin on sleeping and anti-depression
Mice have a habit of being active at night. The authors took advantage of this habit to test the nonRapid Eye Movement (REM) sleep-inducing effects of crocin. The Electroencephalogram (EEG) of mice pre-administered with crocin was measured using an EEG measuring device, and the results showed that non-REM sleep was apparently induced by the administration of at least 10 mg/kg crocin even at 8 pm. The intensity of non-REM sleep increased dose-dependently by the administration of crocin (10-100 mg/ kg). It became also evident that the sleep effects of crocin were not due to inhibition of exercise [34]. As previously indicated the SE prolonged the sleeping time induced by hexobarbital by a sedative effect [35]. These results indicate that crocin and saffron induce good sleep [36,37].
Ohara et al. confirmed that sleep disorders and the concomitant use of hypnotic drugs induced a risk of dementia in aged patients in the Hisayama epidemiological study in Fukuoka, Japan. Compared patients with daily sleep durations shorter than 5.0 h and patients more than 10 h, the risk of dementia for the short sleep duration group increased 2 fold compared with the 10 h group. This evidence suggested that poor sleep may induce brain aging and β-amyloid increase resulting in Alzheimer’s disease. Moreover, sleep disorder may promote inflammation and induce depression and dementia [38]. It is reported that sleep disorder may increases inflammatory processes resulting in Alzheimer’s disease [39]. Several groups indicated that sleep disorder may be an important risk factor for Alzheimer’s disease resulted that it may be an important target for Alzheimer’s disease prevention [40,41]. The connection between sleep disorders and dementia was reported recently [41].
Depression has been considered closely related to early stage dementia and identified as a risk factor for dementia after 2-17 years [42]. In a follow-up of 10,189 patients, depression increased dementia risk later in life in the Whitehall Study in the United Kingdom [43]. Epidemiological study associated with initially cognitive healthy men (4,922) of 71-89 years old during 14 years reported that depression induced a significant incidence of dementia [44].
Since sleeping disorders and depression are closely associated with dementia, the indirect relationship between saffron and crocin and dementia via sleeping disorders or depression was correctly determined.
Clinical studies of saffron on dementia patients
Saffron has been used as herb medicine, spice and pigment for thousands of years and is admitted in Generally Recognized as Safe (GRAS) by the American Food and Drug Administration (FDA) [45]. This commission shows that saffron is safe herb medicine without side effects and can be used in clinical trials as following.
Forty-six Alzheimer's patients were treated with saffron extract 15 mg capsules or placebo twice in a day. After 16 weeks, the saffron group showed clear enhancement in cognitive function. No side effects were observed compared with the case with placebo. The results suggested that 4 weeks administration of saffron extract is effective and safe for mild to moderate Alzheimer's disease patients [46].
A 22-week clinical trial was conducted on 54 patients with Alzheimer's disease using the Cognitive subscale and clinical dementia rating scale. Fifteen mg of saffron extract was administered twice in a day, and 5 mg of donepezil was administered twice daily as a positive control. The results concluded that the saffron extract for 22-weeks administration was effective comparing with donepezil for mild to moderate Alzheimer's disease. Although nausea was occurred in the case of saffron extract as a side effect, mild symptom was observed rather than donepezil. From these evidences, saffron extract can be used as a safe therapeutic drug for Alzheimer's disease [47].
Conclusion
It became evident that crocin ameliorates alcohol-induced inhibition of memory learning and LTP suppression and the inhibitory effects of ethanol via N-methyl-D-aspartate (NMDA) receptors in hippocampal neurons resulting that crocin may improve memory learning based on the above action [48]. Crocin protects brain neurons by inhibiting anti-oxidant and anti-inflammation stress, inhibiting ceramide production, promoting GSH production resulting in inhibiting neuronal cell apoptosis resulting in the prevention of dementia. Saffron is an extremely safe natural medicine that does not cause toxicity even when administered to mice at 5 g/kg [49].
From the above evidences saffron and crocin are considered to be a potential drugs for dementia because they are safe and have various useful activities on the brain.
Acknowledgements: The authors greatly appreciate the Faculty of Pharmacy, Nagasaki International University for providing laboratory facilities and their support of this project. Also we appreciate the Ministry of Agriculture, Forestry and Fishers for the assistance received for the project on measures to promote and revitalise mountain villages.
References
- Bolhassani A. Bioactive Components of Saffron and Their Pharmacological Properties. Stud Nat Prod Chem. 2018; 58: 289-311.
- Morimoto S, Umezaki Y, Shoyama Y, Saito H, Nishi K, et al. Post-harvest degradation of carotenoid glucose esters in saffron. Planta Medica. 1994; 60: 438-440.
- Caballero-Ortega H. Pereda R. HPLC quantification of major active components from 11 different saffron (Crocus sativus L.) sources. Food Chem. 2007; 100: 1126-1131.
- Ordoudi SA, Cagliani LR, Lalou S, Naziri E, Tsimidou MZ, et al. H NMR-based metabolomics of saffron reveals markers for its quality deterioration. Food Res Int. 2015; 70: 1-6.
- Alehosseini A, Gómez-ascaraque LG, Ghorani B, López-Rubio A. Stabilization of a saffron extract through its encapsulation within electrospun/electrosprayed zein structures. LWT. 2019; 113: 108280.
- Zhang Y, Shoyama Y, Sugiura M, Saito H. Effects of Crocus sativus L. on the Ethanol-Induced Impairment of Passive Avoidance Performances in Mice. Biol Pharm Bull. 1994; 17: 217-221.
- Sugiura M, Shoyama Y, Saito H. Crocin (crocetin di-gentiobiose ester) prevents the inhibitory effect of ethanol on long-term potentiation in the dentate gyrus in vivo. J Pharmacol Exp Ther. 1994; 271: 703-707.
- Shimada K, Ishii N, Ohishi K. Structure-activity relationship of cardiac steroids having a double linked sugar and related compounds for the inhibition of Na +, K + -adenosine triphosphatase. J Pharmaco-Dynamics. 1986; 755-759.
- Takemoto T, Ueyama T, Saito H. Potentiation of nerve growth factor-mediated nerve fiber production in organ cultures of chicken embryonic ganglia by ginseng saponins: structure-activity relationship. Chem Pharm Bull. 1984; 32: 3128-3133.
- Abe H, Sakaguchi M, Konishi H. The effects of saikosaponins on biological membranes. 1. The relationship between the structures of saikosaponins and haemolytic activity. Planta Medica. 1978; 34: 160-166.
- Voutquenne L, Lavaud C, Massiot G. Structure activity relationships of haemolytic saponins. Pharma Biol. 2002; 40: 253-262.
- Xuan L, Tanaka H, Xu Y, Shoyama Y. Preparation of monoclonal antibody against crocin and its characterization. Cytotechnology. 1999; 29: 65-70.
- Soeda S, Ochiai T, Shimeno H, Saito H, Abe K, Tanaka H et al. Pharmacological activities of crocin in saffron, J Nat Med. 2007; 61: 102-111.
- Ochiai T, Ohno S, Soeda S, Tanaka H, Shoyama Y, Shimeno H. Crocin prevents the death of rat pheochromyctoma (PC-12) cells by its antioxidant effects stronger than those of α-tocopherol. Neurosci Lett. 2004; 362: 61-64.
- Wang C, Cai X, Hu W, Li Z, Kong F, Chen X, Wang D. Investigation of the neuroprotective effects of crocin via antioxidant activities in HT22 cells and in mice with Alzheimer’s disease. Int J Mol Med. 2018; 956-95
- Ochiai T, Soeda S, Ohno S, Tanaka H, Shoyama Y, Shimeno H. Crocin prevents the death of PC-12 cells through sphingomyelinase-ceramide signaling by increasing glutathione synthesis. Neurochem Int. 2004; 44: 321-330.
- Colombaioni L, Frago LM, Varela-Nieto I, Pesi R. Garcia-Gil M. Serum deprivation increases ceramide levels and induces apoptosis in undifferentiated HN9.10e cells. Neurochemical International. 2002; 49: 327-336.
- Verheij M, Bose R, Lin XH, Yao B, Jarvis WD, Grant S. Requirement for Ceramide-Initiated SAPK/JNK Signalling in StressInduced Apoptosis. Nature. 1996; 380: 75-79.
- Rukenstein A, Rydel RE, Greene LA. Multiple Agents rescue PC12 Cells from Serum-free Cell Death by Translation- and Transcription independent Mechanisms. J Neurosci. 1991; 11: 2552-2563.
- Soeda S, Ochiai T, Paopong L, Tanaka H, Shoyama Y, Shimeno H. Crocin suppresses tumor necrosis factor-induced cell death of neuronally differentiated PC-12 cells. Life Sci. 2001; 69: 2887-2898.
- Sugiura M, Shoyama Y, Saito H, Abe K. The effects of ethanol and crocin on the induction of long-term potentiation in the region of rat hippocampal slices, Jap J Pharmacology. 1995; 67: 395-397.
- Bastani S, Vahedian V, Rashid M, Mir A, Mirzaei S, et al. An evaluation on potential anti-oxidant and anti-inflammatory effects of Crocin. Biomed Pharmacoth. 2022; 153: 113297.
- Zeynab MY, Ali P, Najafi H. Crocin has anti-inflammatory and protective hemia-reperfusion induced renal injuries. Iran J Basic Med Sci. 2017; 20:753-759.
- Yuan Y, Shan X, Men W, Zhai H, Qiao X, et al. The effect of crocin on memory, hippocampal acetylcholine level, and apoptosis in a rat model of cerebral ischemia. Biomed Pharmacother. 2020; 130: 110543.
- Asadi F, Jamshidi AH, Khodagholi F, Yans A, Azimi L, et al. Reversal effects of crocin on amyloid β-induced memory deficit: Modification of autophagy or apoptosis markers. Pharmacol Biochem Behav. 2015; 39: 47-58.
- Du J, Li Y, Song D, Liu J, Huang Q, et al. Protective effects of crocin against endogenous Aβ-induced neurotoxicity in N2a/APP695swe cells. Psychopharmacol (Berl); 2021: 238: 2839-2847.
- Farkhondeh T, Samarghandian S, Yazdi HS, Samini F. The protective effects of crocin in the management of neurodegenerative diseases: a review, Am J Neurodegener Dis. 2018; 7: 1-10.
- Ali A, Yu L, Kousar S, Khalid W, Maqbool Z, Aziz A. Crocin: Functional characteristics, extraction, food applications and efficacy against brain related disorders. Front Nutr. 2022.
- Papandreou MA, Kanakis CD, Polissiou MG, Efthimiopoulos S, Cordopatis P, Margarity M. Inhibitory Activity on Amyloid-Beta Aggregation and Antioxidant Properties of Crocus Sativus Stigmas Extract and its Crocin Constituents. J Agricul Food Inform. 2006; 54: 8762–8768.
- Roustazade R, Radahmadi M, Yazdani Y. Therapeutic Effects of Saffron Extract on Different Memory Types, Anxiety, and Hippocampal BDNF and TNF-α Gene Expressions in Sub-chronically Stressed Rats. Nutr Neurosci. 2021; 25: 192-206.
- Zhang Y, Shoyama Y, Sugiura M, Saito H. Effects of Crocus sativus L. on the Ethanol-Induced Impairment of Passive Avoidance Performances in Mice. Biolog Pharmaceut Bull. 1994; 17: 217-221.
- Sugiura M, Shoyama Y, Saito H. Crocin improves the ethanol-inducd impairment of leaning behaviors and mice in in passive avoidance tasks. Proc J Acad Ser B: Phys Biolog Sci. 1995; 71: 319-324.
- Sugiura M, Shoyama Y, Saito H, Abe K. Ethanol extract of Crocus sativus L. antagonizes the inhibitory action of ethanol on hippocampal long-term potentiation in vivo. Phytoth Res. 1995; 9: 100-104.
- Masaki M, Aritake K, Tanaka H, Shoyama Y, Huang ZL, et al. Crocin promotes non-rapid eye movement sleep in mice. Mol Nutr Food Res. 2012; 56: 304-308.
- Zhang Y, Shoyama Y, Sugiura M, Saito H. Effects of Crocus sativus L. on the Ethanol-Induced Impairment of Passive Avoidance Performances in Mice. Biol Pharm Bull. 1994; 17: 217-221.
- Takeda R, Yamamoto K, Tokuda M, Koga J, Higaki M, et al. Improvement in quality of sleep by the intake of saffron derived crocin and safranal-A randomized, double blind, placebo controlled, parallel group trial. Jap Pharmacol Therap. 2020; 48: 497-504.
- Nishida A, Fujita T, Nagaregawa Y, Shoyama Y, Ohnuki K, et al. Sleep enhancement by saffron extract affron in randomized control trial. Jap Pharmacol Therap. 2018, 46. 1407-1415.
- Ohara T, Honda T, Hata J, Yoshida D, Mukai N, et al. Association between Daily Sleep Duration and Risk of Dementia and Mortality in a Japanese Community. J Am Geriatr Soc. 2018; 66: 1911-1918.
- Irwin MR, Vitiello MV. Implications of Sleep Disturbance and Inflammation for Alzheimer’s Disease Dementia. Lancet Neurol. 2019; 18: 296-306.
- Bubu OM, Brannick M, Mortimer J, Umasabor-Bubu O, Sebastião YV, et al. Sleep, Cognitive Impairment, and Alzheimer’s Disease: A Systematic Review and Meta-Analysis. Sleep. 2017; 40: 1 Jan.
- Livingston G, Huntley J, Sommerlad A, Ames D, Ballard C, et al. Dementia Prevention, Intervention, and Care: 2020 Report of the Lancet Commission. Lancet. 2020; 396: 413-446.
- Prince M, Albanese E, Guerchet M, Prina M. World Alzheimer Report 2014 - Dementia and Risk Reduction: An Analysis of Protective and Modifiable Risk Factors. London, UK: Alzheimer’s Disease International. 2014.
- Almeida OP, Hankey GJ, Yeap BB, Golledge J, Flicker L. Depression as a Modifiable Factor to Decrease the Risk of Dementia. Transl Psychiatry. 2017; 7: e1117.
- Kelly M, Duff H, Kelly S, Power JEM, Brennan S, et al. The Impact of Social Activities, Social Networks, Social Support and Social Relationships on the Cognitive Functioning of Healthy Older Adults: a Systematic Review. Syst Rev. 2017; 6: 259.
- Department of Health and Human Services; Subchapter B - Food For Human Consumption (Continued); Part 182.10 - Substances Generally Recognized as Safe (GRAS).
- Saeedi M, Rashidy-Pour A. Association between chronic stress and Alzheimer’s disease: Therapeutic effects of Saffron. Biomed Pharmacoth. 2021; 133: 110995.
- Akhondzadeh S, Shafiee-Sabet M, Harirchian MH. A 22-week, multicenter, randomized, double-blind controlled trial of Crocus sativus in the treatment of mild to-moderate Alzheimer’s disease. Psychopharmacology (Berl). 2010; 207: 637-643.
- Abe K, Sugiura M, Shoyama Y, Saito H. Crocin Antagonizes Ethanol Inhibition of NMDA Receptor-Mediated Responses in Rat Hippocampal Neurons. Brain Res. 1998; 787: 132-138.
- Lymperopoulou CD, Lamari FN. Saffron safety in humans: lessons from the animal and clinical studies. Med Aromat Plants. 2015; 4: e164.
