
Open Access, Volume 9
A Universal approach to cell separation based on reversible biotin analog with streptavidin magnetic beads
Laiqiang Ying1*; Andy Ying2
1ABP Biosciences 405 E Gude Dr, Rockville, MD 20850, USA.
2Montgomery Blair High School 51 University Blvd E, Silver Spring, MD 20901, USA.
Laiqiang Ying
ABP Biosciences 405 E Gude Dr, Rockville, MD 20850, USA.
Email: lqying@abpbio.com
Received : July 22, 2023,
Accepted : Sep 04, 2023
Published : Sep 08, 2023,
Archived : www.jclinmedcasereports.com
Abstract
Cell separation is a critical process in biological and biomedical research, and in clinical therapy. The gold standard of cell separation techniques are fluorescence-activated cell sorting (FACS) and magneticactivated cell sorting (MACS). Despite this, there are still some limitations with FACS and MACS such as low recovery and purity, and affecting the cell physiology and viability. Herein, we report a universal cell separation approach based on reversible binding between N3′-ethyl biotin and streptavidin, which allows for fast capturing of target cells from cell mixture, and fast and complete releasing cells from magnetic beads under mild, physiological conditions. This universal method for detaching cells from magnetic beads provides an efficient cell separation technique based on MACS to give high recovery, high purity, and high viability of cells.
Keywords: Cell separation; Biotin; Streptavidin; Immunomagnetic bead; Immobilization.
Copy right Statement: Content published in the journal follows Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0). © Ying L (2023)
Journal: Open Journal of Clinical and Medical Case Reports is an international, open access, peer reviewed Journal mainly focused exclusively on the medical and clinical case reports.
Citation: Ying L, Ying A. A universal approach to cell separation based on reversible biotin analog with streptavidin magnetic beads. Open J Clin Med Case Rep. 2023; 2107.
Short Commentary
Cell separation is a powerful tool enabling the isolation of cellular subpopulations for biological research and clinical applications [1-5]. Many cell separation techniques are currently commercially available, which are predominantly based on three methodologies: adherence, density, and cell surface marker. Some new techniques based on microfluidic devices and cellular properties are being developed [6,7]. The gold standard for cell separation is fluorescence-activated cell sorting (FACS) and magnetic-activated cell sorting (MACS) [8-11]. These techniques have been extensively used in many applications ranging from in vitro diagnostics to cell-based therapeutics [12-18]. FACS is capable of processing millions of cells and isolating multiple, high purity subpopulations. However, the limitations of this technology are the timeconsuming, high cost, and low recovery, especially in the separation of rare cells [19,20]. MACS is designed for high-throughput separation of specific target cells from complex mixtures based on immuno-magnetic labels [11,21]. These MACS techniques have been classified based on the size of magnetic particles, such as MACS® (20-100 nm, Miltenyi Biotec); IMAG® (100-500 nm, BD Biosciences); Dynal® (1-5 µm, ThermoFisher Scientific). The MACS® technique usually doesn’t require the bead removal step after positive cell separation, due to the small size (~50 nm) and biodegradability of magnetic particles [22]. But, a strong external magnetic filed must be applied to capture the small size of magnetic particles. In some particular applications such as cell sequential sorting based on more than one surface marker (MultiSort strategy), the prior removal of the beads used in the first sorting step is required for next-step selection [23,24]. Using microbeads such as Dynabeads (1-5 µm) for cell separation allows for being easily “pulled out” from cell suspension with a simple magnetic separator such as an MPC separator [25,26]. However, in many cases, the removal of larger superparamagnetic particles from the cell surface is necessary for down-stream applications [11].
To detach magnetic beads from the cell surface, several approaches have been developed [11]. For example, chymopapain has been used to selectively cleave the CD34 molecule from human hematopoietic cells to detach magnetic beads coated with anti-CD34 antibody [27]. But, this method is only useful for CD34+ cell separation. For general applicability, trypsin has been used for proteolytic detachment [28]. However, trypsin digestion is non-specific, and thus affects the surface antigen expression of isolated cells. Two methods based on the competition approach have been commercialized. One is called the “DETACHaBEAD” system [29], in which a polyclonal antibody is used to compete with the binding of primary antibody on magnetic beads; The other is called Bartex Isolex system [30], in which a synthetic peptide is used to compete with the binding of the anti-CD34 antibody. A DNA linker between primary antibody and magnetic beads was also used to detach cells from magnetic beads using low-ionic-strength buffer or DNase [31,32]. Overall, all the detach methods have disadvantages in low recovery or requirement of long incubation time, thus affecting the cell physiology and viability [33,34].
Developing a new universal method with fast and complete detachment will be valuable for cell separation applications. In the past, we synthesized a library of new biotin analogs, in which some key H-bonding interactions between biotin and streptavidin were blocked or perturbed to reduce the binding affinity [35,36]. We have identified some biotin analogs that show optimal reversible interaction with streptavidin, which allows (1) fast and complete binding and (2) fast and complete release, under mild, physiological conditions. As shown in Figure 1, one of our new biotin analogs, named N3′-ethyl biotin (kD ~0.8 nM), has fast on-rate (kon ~1.5 x 105 M-1S-1), and also fast off-rate (koff ~5.8 x 10-2 S-1) in the presence of 1 mM bisbiotin competing reagent. Instead, biotin is almost irreversible under the same condition. These reversible interactions between N3′-ethyl biotin and streptavidin provide a unique method for reversible immobilization of N3′-ethyl biotinylated antibody on magnetic streptavidin beads for cell separation applications. Here we report a universal cell separation approach based on reversible binding between N3′-ethyl biotin and streptavidin.
To assess the effectiveness of cell separation using the streptavidin-N3′-ethyl biotin system, the N3′- ethyl biotin was initially labeled with mouse anti-human CDx monoclonal antibodies (mAb). In addition, the mAbs were conjugated with AlexaFluor 488 to allow for flow cytometry analysis. The evaluation of cell separation was performed using a cell model system of human peripheral T lymphocytes (CD3+, CD4+, and CD8+ cells). The human peripheral blood mononuclear cells (PBMCs) were prepared by NH4Cl lysis from human peripheral blood obtained from healthy volunteers following company guidelines. Initially, PBMCs (3 x 107 cells) were separately incubated with mouse anti-human CDx (CD3, CD4, and CD8) mAb-N3′-ethyl biotin conjugate for 10 minutes on ice for 10 min. Then, streptavidin M280 Dynabeads (ThermoFisher Scientific) were added and incubated under rolling and tilting for 10 minutes at room temperature. The cell mixture was then placed on a magnet for 1 minute, and the supernatant was aspirated. The cells containing magnetic beads were washed, and then the cells were detached from the magnetic beads by incubating with 1 mM bis-biotin for 5 minutes. The number and viability of isolated cells were measured using Countess® automated cell counter. The purity and depletion efficiency was evaluated using flow cytometry. The results are summarized in Figure 2 and Table 1. By demonstrating the feasibility of cell separation based on three different cell surface markers, a simple workflow has been established, consisting of two-step incubations, one-step wash, and one-step detachment, as shown in Figure 2a. The entire cell separation process can be carried out in less than 30 min to achieve high recovery (>80%), high purity (>92%), and high viability (>92%). By using streptavidin M280 Dynalbeads, each target cell was captured by multiple magnetic beads (Figure 2b), and can be easily separated by a simple magnet (Figure 2b) to achieve high depletion efficiency (98-100%, Figure 2c). The N3′-ethyl biotinylated mAb was rapidly and fully released from the streptavidin magnetic beads using 1 mM bis-biotin under physiological conditions, resulting in high recovery, high purity, and high viability of cells (Figure 2c).
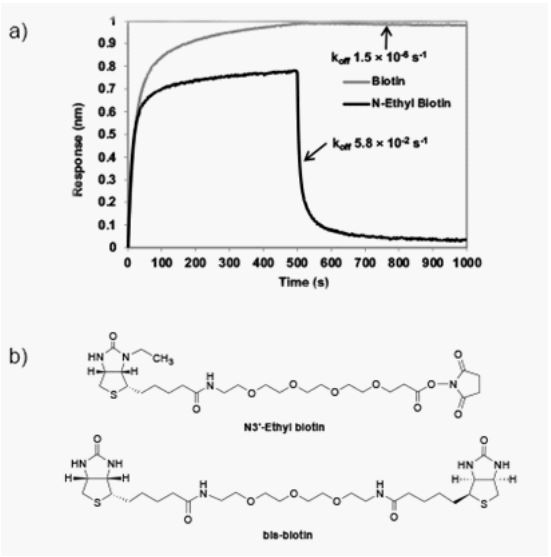
Figure 1: a) Association and dissociation curves between biotin or N3′-ethyl biotin and
streptavidin on the ForteBio system. Each association curve was generated by incubating
biotin or N3′-ethyl biotin sensor tip with streptavidin. The dissociation curve was generated by incubating the biotin-streptavidin or N3′-ethyl biotin-streptavidin complex at 1
mM bis-biotin in the PBS buffer. b) Chemical structures of N3′-ethyl biotin and bis-biotin.
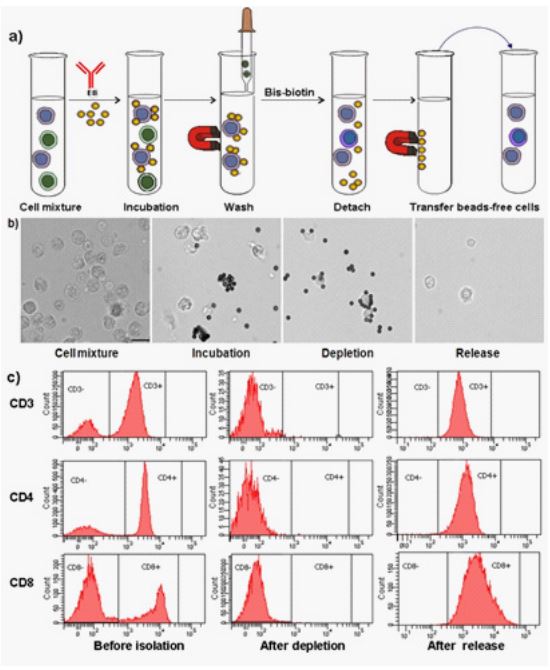
Figure 2: a) Illustration of cell separation workflow; b) Cell imaging at different separation stages. Scale bar, 10 µm; c) Flow histograms of cells based on three different cell
surface markers (CD3, CD4, and CD8) at pre-isolation, after depletion, and after release,
respectively.
The method as described above using primary antibody with streptavidin M280 Dynabeads requires the labeling of each primary antibody (anti-CD3 mAb, anti-CD4 mAb, and anti-CD8 mAb) with N3′-ethyl biotin molecule. The process of antibody labeling can be time-consuming if the cell separation involves various cell surface makers such as CD1 to CD200. To explore a more general and cost-effective method, a secondary antibody such as goat anti-mouse (GAM) IgG was labeled with N3′-ethyl biotin. Then, the N3′- ethyl biotin-GAM IgG conjugate was immobilized onto streptavidin M280 Dynabeads. The same procedure as above was followed for cell separations except for the use of a secondary antibody to label N3′-ethyl biotin. Briefly, PBMCs (3 x 107 cells) were incubated with unlabelled mouse anti-human CDx (CD3, CD4, and CD8) mAbs on ice for 10 minutes. Streptavidin M280 Dynabeads coated with GAM IgG-N3′-ethyl biotin conjugate was added and incubated under rolling and tilting for 10 minutes at room temperature. After washing to remove unbound reagents, the magnetic beads were detached from cells by incubating with 1 mM bis-biotin for 5 minutes to recover target cells free of beads. The results are shown in Table 1. By conjugating a secondary antibody such as GAM IgG with N3′-ethyl biotin, it eliminates multiple conjugation processes on primary antibodies such as mouse anti-human CDx mAb labeling. This method provides a more cost-effective and universal approach. As demonstrated in Table 1, the recovery and purity of isolated cells using the secondary antibody approach are better than those using the primary antibody approach. This could be due to the higher affinity of the excess amount of secondary antibody-coated streptavidin magnetic beads to capture primary antibody-labeled targeted cells, and it tolerates more extensive wash to achieve high purity. In summary, both approaches provide a simple workflow and a universal detachment method for cell separation applications.
Table 1: Recovery, purity, and viability of isolated cells.
| Method | Cell type | Recovery[a] | Purity[b] | Viability[a] |
|---|---|---|---|---|
| Primary Ab | CD3 | 87 ± 6% | 96 ± 2% | 96 ± 2% |
| Primary Ab | CD4 | 84 ± 8% | 92 ± 2% | 92 ± 3% |
| Primary Ab | CD8 | 72 ± 5% | 93 ± 3% | 93 ± 2% |
| Secondary Ab | CD3 | 92 ± 4% | 96 ± 2% | 97 ± 2% |
| Secondary Ab | CD4 | 88 ± 6% | 97 ± 2% | 94 ± 2% |
| Secondary Ab | CD8 | 82 ± 5% | 96 ± 2% | 95 ± 3% |
[a] Each recovery and viability are reported the mean of independent triplicate experiments based on Countess® measurement.
[b] Each purity is reported the mean of independent triplicate experiments based on flow cytometry measurement. Error bars
represent the standard deviation of independent triplicate experiments.
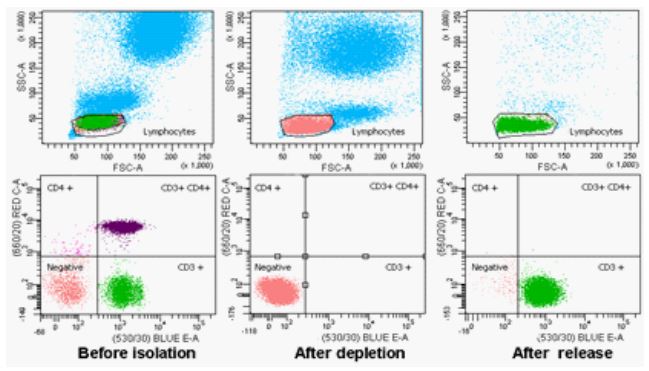
Figure 3: Flow histograms of cells based on dual cell surface markers (CD3 and CD4) at
pre-isolation, after depletion, and after release. The population of lymphocytes, monocytes, and granulocytes is shown on top. The subpopulation of lymphocytes based on CD3
and CD4 cell surface markers is shown at the bottom.
An important application in cell separation is to isolate cell subpopulations based on dual cell surface markers. Miltenyi Biotec has developed a MultiSort strategy, which involves first incubating a cell mixture with MultiSort MicroBeads to capture cells of interest based on the first cell surface marker, and then enzymatically cleaving the MultiSort MicroBeads. The isolated cells are then incubated with MACS® MicroBeads to capture the cell subset based on the second cell surface marker. Another method of cell separation based on dual cell surface markers involves using two different sizes of immunomagnetic beads (MACS® MicroBeads and M280 Dynabeads), which eliminates the need for detaching MACS® MicroBeads used in the first selection step [37]. However, both strategies require a two-step sequential selection. To develop a more straightforward workflow of cell separation based on dual cell surface markers, we proposed a new approach using biotin and biotin analog with different binding affinity. To test the hypothesis, we chose a cell model system of human peripheral T lymphocytes based on dual cell surface markers (CD3 and CD4). The anti-CD4 mAb was labeled with biotin molecule and AlexaFluor 647 (Labeling AlexaFluor dye is for flow cytometry analysis purposes); and anti-CD3 mAb was labeled with N3′-ethyl biotin and AlexaFluor 488. PBMCs (3 × 107 cells) were incubated with anti-CD4 mAb-biotin conjugate and anti-CD3 mAb-N3′- ethyl biotin conjugate simultaneously on ice for 10 minutes. Then, streptavidin M280 Dynabeads were added and incubated for 10 minutes at room temperature. After the washing step to remove unbound cells, the magnetically labeled cells were selectively released by incubating with 1 mM bis-biotin for 5 minutes to yield beads-free CD3+CD4− cells. As shown in Figure 3, after incubation with biotinylated anti-CD4 mAb and N3′-ethyl biotinylated anti-CD3 mAb, the cells either CD3+ or CD4+ have been captured by streptavidin M280 Dynabeads with depletion efficiency (~100%). After incubating with 1 mM bis-biotin, CD3+CD4− cells were selectively detached from magnetic beads with 80 ± 5% recovery, 92 ± 3% purity and 95 ± 2% viability, and CD4+ cells remain bound to magnetic beads. These results show that the N3′-ethyl biotin can be selectively detached from streptavidin without interfering with biotin-streptavidin interactions by controlling the release conditions. This strategy provides a simple one-step cell selection method based on dual cell surface markers.
Conclusions
In summary, a universal approach to cell separation has been developed using the reversible N3′- ethyl biotin-streptavidin system. The reversible binding of N3′-ethyl biotin to streptavidin allows for the efficient binding of target cells to streptavidin beads and the complete detachment of cells from streptavidin beads under physiological conditions. To the best of our knowledge, the N3′-ethyl biotin-streptavidin system represents the best universal method for gentle capture and release. This system provides a new type of cell separation platform that holds promise for clinical diagnostics and cell-based therapy.
References
- KT. Guo, R. Schafer, A. Paul, A. Gerber, G. Ziemer, H. P. Wendel, Stem Cells 2006, 24, 2220-2231.
- S Takaishi, T Okumura, S Tu, SW Wang, W Shibata, et al. Stem Cells. 2009; 27: 1006-1020.
- VH Terry, ICD Johnston, CA Spina, Virology. 2009; 388: 294-304.
- R Handgretinger, P Lang, M Schumm, G Taylor, S Neu, et al. Bone Marrow Transplant. 1998; 21: 987-993.
- G Mancardi, R Saccardi. Lancet Neurol. 2008; 7: 626-636.
- F. Petersson, L. Åberg, A. M. Swärd-Nilsson, T. Laurell, Anal. Chem. 2007; 79: 5117-5123.
- X Hu, PH Bessette, J Qian, CD Meinhart, PS Daugherty, et al. Proc. Natl. Acad. Sci. USA. 2005; 102: 15757-15761.
- LA Herzenberg, D Parks, B Sahaf, O Perez, M Roederer. Clin. Chem. 2002; 48: 1819-1827.
- S Miltenyi, W Muller, W Weichel, A Radbruch. Cytometry. 1990; 11: 231-238.
- TT Hansel, IJ De Vries, T Iff, S Rihs, M Wandzilak, et al. J. Immunol. Methods. 1991; 145: 105-110.
- I Safarik, M Safarikova, J Chromatogr. B Biomed. Sci. Appl. 1999; 722: 33-53.
- BK Patterson, M Till, P Otto, C Goolsby, MR Furtado, et al. Science. 1993; 260: 976-979.
- O Olsvik, T Popovic, E Skjerve, KS Cudjoe, E Hornes, et al. Clin. Microbiol. Rev. 1994; 7: 43-54.
- L Battistini, L Piccio, B Rossi, S Bach, S Galgani, et al. Blood. 2003; 101: 4775-4782.
- NB Ivanova, JT Dimos, C Schaniel, JA Hackney, KA Moore, et al. Science. 2002; 298: 601-604.
- E Thomas, R Storb, RA Clift, A Fefer, FL Johnson, et al. N. Engl. J. Med. 1975; 292: 832-843.
- ED Thomas, R Storb, RA Clift, A Fefer, L Johnson, et al. N. Engl. J. Med. 1975; 292: 895-902.
- M Muller, BK Fleischmann, S Selbert, GJ Ji, E Endl, et al. FASEB J. 2000; 14: 2540-2548.
- SF Ibrahim, G van den Engh. Curr. Opin. Biotechnol. 2003; 14: 5-12.
- BA Sutermaster, EM Darling. Sci. Rep. 2019; 9: 227.
- A Grutzkau, A Radbruch. Cytometry A. 2010; 77: 643-647.
- B Schmitz, A Radbruch, T Kummel, C Wickenhauser, H Korb, et al. Eur. J. Haematol. 1994; 52: 267-275.
- J Busch, P Huber, E Pfluger, S Miltenyi, J Holtz, et al. Prenat. Diagn. 1994; 14: 1129-1140.
- XX Zhao, Y Ozaki, N Suzumori, T Sato, K Suzumori. Congenit. Anom. (Kyoto) 2002; 42: 120-124.
- PW Hewett, JC Murray. Eur. J. Cell. Biol. 1993; 62: 451-454.
- CJ Jackson, PK Garbett, B Nissen, L Schrieber. J. Cell. Sci. 1990; 96: 257-262.
- F Silvestri, S Banavali, M Yin, V Gopal, C Savignano, et al. H. D. Preisler, Haematologica. 1992; 77: 307-310.
- P Collin-Osdoby, MJ Oursler, D Webber, P Osdoby. J. Bone Miner Res. 1991; 6: 1353-1365.
- G Anderson, EJ Jenkinson, NC Moore, JJ Owen, Nature. 1993; 362: 70-73.
- SD Rowley, M Loken, J Radich, LA Kunkle, BJ Mills, et al. Bone Marrow Transplant. 1998; 21: 1253-1262.
- WH Scouten, P Konecny. Anal. Biochem. 1992; 205: 313-318.
- C Ljungquist, J Lundeberg, AM Rasmussen, E Hornes, M Uhlen. DNA Cell Biol. 1993; 12: 191-197.
- J Seidl, R Knuechel, LA Kunz-Schughart. Cytometry. 1999; 36: 102-111.
- A Tiwari, G Punshon, A Kidane, G Hamilton, AM Seifalian. Cell Biol. Toxicol. 2003; 19: 265-272.
- L Ying, BP Branchaud, Y Zhang, S Yue. US9567346B2. 2017.
- L Ying, BP Branchaud. Chem. Commun. 2011; 47: 8593-8595.
- KM Partington, EJ Jenkinson, G Anderson, J. Immunol. Methods. 1999; 223: 195-205.
