
Open Access, Volume 9
Identification of DNA repair gene signature and potential molecular subtypes in hepatocellular carcinoma
Yutao Wang*
Department of Urology, China Medical University, The First Hospital of China Medical University, Shenyang, China.
Email: 809448033@qq.com
Received : February 02, 2023,
Accepted : March 02, 2023
Published : March 10, 2023,
Archived : www.jclinmedcasereports.com
Abstract
DNA repair is a critical factor in tumor progression since it influences tumor mutational burden, genome stability, PD-L1 expression, tumor-infiltrating lymphocytes (TILs) and immunotherapy response. In this study, we constructed a prognostic model for hepatocellular carcinoma (HCC) based on DNA damage response (DDR)-related genes. The patients were stratified on the basis of the risk score, and the low-risk group showed better survival rates compared to the high-risk counterparts. The risk model showed good predictive accuracy for 1-, 3- and 5-year survival as per receiver operator curve analysis (ROC). In addition, the risk model was verified as an independent prognostic factor of HCC along with tumor stage. We further constructed a nomogram based on the independent factors for predicting overall survival. Gene set enrichment analysis (GSEA) demonstrated that cell cycle, apoptosis, MAPK, mTOR and the WNT cascades were enriched in the high-risk group. Two potential molecular subtypes of HCC were identified based on the expression of DDR genes in the training and validation datasets. The two subtypes differed in terms of immune cell infiltration and expression of immune checkpoint receptors. Taken together, we identified potential biomarkers of HCC prognosis that may provide novel insights into the molecular mechanisms underlying HCC based on cell assays.
Keywords: Hepatocellular carcinoma; DNA repair gene; Prognosis; Molecular subtypes of HCC; Immune infiltrition.
Copy right Statement: Content published in the journal follows Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0). © Wang Y (2023)
Journal: Open Journal of Clinical and Medical Case Reports is an international, open access, peer reviewed Journal mainly focused exclusively on the medical and clinical case reports.
Citation: Wang Y. Identification of DNA repair gene signature and potential molecular subtypes in hepatocellular carcinoma. Open J Clin Med Case Rep. 2022; 1991.
Introduction
According to the 2018 global cancer statistics, hepatocellular carcinoma (HCC) is the sixth most common malignancy and the fourth leading cause of cancer-related mortality [1,2]. Despite recent advances in HCC treatment, the outcomes are far from satisfactory [3,4]. Therefore, there is an urgent need to identify novel therapeutic targets and diagnostic biomarkers of HCC in order to improve patient prognosis.
Given that DNA damage is a hallmark of cancer cells, the pathways involved in DNA damage and repair are repositories of novel therapeutic targets [5]. Studies show that genes involved in DNA damage response (DDR) pathways such as base excision repair, mismatch repair and nucleotide excision repair [6] are aberrantly expressed during cancer development and progression [7-10]. Dysregulated DDR is associated with increased genome instability in HCC cells [11] and is therefore prognostically significant.
In recent years, with the rise of high-throughput sequencing technology, sequencing data and clinical follow-up information in many cancer databases can be downloaded by us. We therefore attempted to download the hepatocyte dataset from TCGA and GEO databases. At present, many studies have used prognostic model methods to predict the overall survival rate of patients, and compared with clinical indicators such as clinical stage and pathological stage, it has shown excellent superiority.
In this study, we examined the prognostic potential of DDR-linked genes in HCC and developed a risk model. To this end, 150 DDR-related genes were identified from the MSigDB database, and an 11-gene HCC prognostic signature was constructed following univariate cox regression and random forest analyses. The robustness of the model was ascertained by internal and external validation. The potential pathways in HCC associated with the risk model were also identified by GSEA, and the correlation between clinical traits and the risk score was analyzed. Finally, we identified and validated two molecular subtypes of HCC based on DDR gene expression. Taken together, our findings provide novel insights into the molecular mechanism of HCC and establish an independent DDR gene-based prognostic signature.
Methods
Data collection: Gene expression and clinical data of HCC samples were obtained from TCGA-LIHC (https://portal.gdc.cancer.gov/) and ICGC-LIRI (https://dcc.icgc.org/) datasets. DDR-linked genes were retrieved from MSigDB, V7.1 (https://www.gsea-msigdb.org/gsea/msigdb), and those present in both datasets were retained.
Risk signature construction: The prognostic DDR-linked genes were identified in the LIHC and LIRI-JP datasets through univariate and multivariate Cox regression analyses. The risk score was calculated for each patient in both datasets as follows: Risk score = (Exp i*β i), where Expi is the expression level of prognostic genes and β i is the cox regression coefficient for each prognostic gene. The patients were classified into high- and low-risk groups according to the median risk score. The “survival” R package and “survminer” package were used to examine survival differences between the high- and low-risk groups. The predictive accuracy of the risk model for 1-, 3- and 45-year survival was evaluated using the “survivalROC” package in R (https://cran.r-project.org/web/packages/survivalROC/index.html). Prognostic independence of the gene risk score and clinical features was determined by univariate and multivariate cox regression analyses. Pathways potentially linked to the high- and low-risk groups were identified using GSEA, with c2.cgp.v7.1.symbols.gmt selected as reference gene set.
Nomogram and DCA curve construction: A nomogram was developed based on all independent prognostic factors. The discriminative ability of the nomogram was assessed through a calibration plot using the bootstrap approach with 1,000 replications [12]. Decision curve analysis was performed to determine the benefit of the prognostic factor [13].
Consensus clustering: To determine the potential molecular subtypes of HCC based on the DDR-related genes, the patients were grouped into diverse clusters using “ConsensusClusterPlus” R package (50 iterations, 80% resampling samples) [14]. Principal Components Analysis (PCA) was used to distinguish various LIHC subgroups and all analyses were validated on the LIRI-JP dataset.
Immune infiltration analysis: The ssGSEA algorithm in “GSVA” R package was used to assess the infiltration levels of 28 immune cells in the high- and low-risk groups, and the enrich score of the immune cells in each sample were calculated [15-17]. The expression of immune checkpoint genes in both groups was also analyzed.
Cell culture and transfection: Human HCC cell lines Hep G2 and MHCC-97H were purchased from (National collection of authenticated cell culture, Shanghai, CN), and incubated at 37oC with 5% CO2 in a humidity saturated environment. Cells were cultured in DMED (Hyclone, LA, USA) and supplied with 10% fetal bovine serum (BI, Israel), anti biotics (0.1 U/l penicillin and 100 g/l streptomycin). DGUOK siRNA were obtained from RiboBio Co., Ltd. (Guangzhou, China). The siRNA was dissolved in DEPC-treated water. Lipofectamine 2000 reagent (Invitrogen, CA, USA) were used for transfection according to the manufacturer’s protocol. The solutions were mixed together and incubated at room temperature for 30 minutes. 30 nM siRNA was added into each well and incubated at 37oC.
Hoechst staining: Cell apoptosis was observed by the morphological changes of the cell nucleus (chromatin agglutination or DNA fragmentation). Cells were treated with si-NC or si-DGUOK, and washed with PBS twice, Hoechst 33258 (1 μg/ml) was added for 20 min at room temperature avoiding light. Images were gathered by fluorescence microscope (Nikon, Japan).
Cell viability assays: Cells were seeded in 96-well plates at 10,000 cells per well, and cultured for 24 h. They were treated with si-NC or si-DGUOK. Then CCK-8 were added to each plate, absorbance was measured at 450 nm using a FLUOstar Omega microplate reader (BMG Labtech). Cell viability of samples was calculated according to the manufacturer’s instructions.
Statistical analyses: Statistical analyses were conducted using R (https://www.r-project.org/). Survival analysis was performed using the Kaplan-Meier (KM) method. P<0.05 was considered statistically significant.
Results
Identification of survival-related DDR risk model: The expression data of 150 DDR-related genes was downloaded from TGCA dataset consisting of 343 HCC samples. Univariate cox regression analysis identified 37 prognostic genes associated with the survival of HCC patients. Following stepwise multivariate cox regression analysis, 11 genes including AAAS, CANT1, CLP1, DGUOK, GTF2B, GTF2H1, NCBP2, POLA1, POLE4, POLR2D and POLR2E were retrieved, and used to develop a risk model. The risk score for each patient was computed as follows: AAAS*-0.022 + CANT1*-0.016 + CLP1*-0.098 + DGUOK*-0.016 + GTF2B*0.018 + GTF2H1*0.034 + NCBP2*0.042 + POLA1*0.089 + POLE4*0.015 + POLR2* 0.047 + POLR2E*0.007. On the basis of the median risk score, the patients were classified into the high- and low-risk groups. As shown in Figure 1A, patients in the low-risk group experienced longer survival relative to those in the high-risk group. Furthermore, KM analysis confirmed better prognosis of the low-risk group relative to that of the high-risk group (p value <0.001) (Figure 1B). ROC analysis was performed to determine the predictive performance of the risk model. The area under curve (AUC) values for 1- and 3-year survival were 0.76 and 0.66 respectively (Figure 1C), indicating good accuracy.

Figure 1: Survival analysis of DDR genes in TCGA dataset.
(A) Risk score plot for the DDR signature. Upper panel
shows risk score distribution, middle panel shows case distribution, and the lower panel indicates the expression level
of the 11 DDR genes. (B) KM survival curves of the high-and
low-risk groups. (C) ROC curve analysis of the risk gene signature.

Figure 2: Survival analysis of DDR genes in ICGC dataset.
(A) Risk score plot for the DDR signature. Upper panel
shows risk score distribution. Middle panel shows case distribution. Lower panel shows the expression level of the 11
DDR genes. (B) KM survival curves of high- and low-risk
groups. (C) ROC curve analysis for the risk gene signature.
External validation of the DDR-gene prognostic signature: To assess the reliability and robustness of the 11-gene signature, we downloaded a dataset of 231 HCC samples from ICGC (https://dcc.icgc.org/). The risk scores were calculated for each patient, and the cohort was stratified into high- or low-risk groups. Consistent with the findings above, most surviving cases were classified into the low-risk group, whereas the high-risk group had a higher mortality rate (Figure 2A). KM analysis revealed that the low-risk group patients had better overall survival relative to their high-risk counterparts (Figure 2B). The AUC values for 1- and 3-year survival were 0.73 and 0.77 respectively, suggesting good prognostic performance of the risk model in HCC (Figure 2C).
The risk model is an independent prognostic predictor of HCC: Univariate and multivariate cox regression analyses showed that the risk model and tumor stage were independent risk factors for HCC (Figure 3A-B). In addition, ROC assessment showed that the risk model outperformed tumor stage in predicting 1-year prognosis (AUC 0.746 VS 0.700) (Figure 4A). We then constructed a nomogram to predict overall survival (OS) at 1-, 3- and 5-years based on the risk model and tumor stage (Figure 5A). As shown in Figure 4B-D, the AUC value of the nomogram indicated good prognostic performance at 1-, 3- and 5-years. Furthermore, calibration curve plots validated the stability of the nomogram (Figure 5B-D). Taken together, the nomogram consisting of DDR gene-based risk score and tumor stage can robustly predict the prognosis of HCC patients, and therefore can aid in clinical decision-making.

Figure 4: ROC curve analysis of the nomogram, gene signature and disease stage for 1- (A), 3- (B),
and 5-year (C) survival.

Figure 3: (A) Univariate and (B) multivariate cox regression
analyses demonstrating the prognostic value of the gene biosignature and clinical traits.
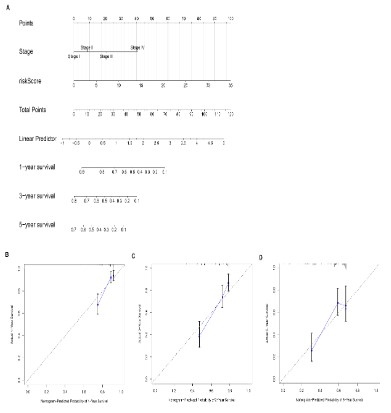
Figure 5: Construction and validation of a prognostic nomogram. (A) A nomogram was developed based on the
risk model and tumor stage to estimate overall survival of
HCC patients. Calibration curve plot of the nomogram for
estimating 1- (B), 3- (C) and 5-year (D) survival of HCC
patients.
Gene Set Enrichment Analysis (GSEA): GSEA was performed for the high-risk and low-risk groups to identify pathways that are significantly enriched in HCC. Pathways related to apoptosis, cell cycle, and MAPK, mTOR, NOCTH, UBIQUITIN and WNT signaling were enriched in the high-risk group, whereas the low-risk group showed significant enrichment of fatty acid metabolism and retinol metabolism pathways (Figure 6A-B). Thus, the low-risk score and favorable prognosis are correlated with metabolism-linked cascades, whereas cancer-related pathways coincide with poor prognosis and high-risk score.

Figure 6: Gene set enrichment analysis results showing enriched pathways in (A) high-risk and (B) low-risk groups.
Identification of molecular subtypes of HCC: The consensus clustering algorithm was used to identify molecular subtypes of HCC on the basis of the DDR genes. Based on the cumulative distribution function curve and the consensus heatmap, we set K = 2 as the optimal cluster (Figure 7A-C). PCA further demarcated the patients into two distinct subgroups (Figure 7D), and the overall survival in subgroup 1 was better than in subgroup 2 (Figure 7E). In addition, we validated the subgroups in the ICGC dataset, which confirmed the robustness of the classification (Figure 8). The correlation between the subgroups and the clinical characteristics in TCGA and ICGC datasets revealed that the group with better survival outcomes included more early-stage cases (Figure 9A-B).
Correlation of the immune infiltration with HCC subclasses: The ssGSEA algorithm were used to analyze the infiltration of 28 immune cells in the high- and low-risk groups. The infiltration levels of activated CD4+ T cells, central memory CD4+ T cells, central memory CD8+ T cells, effector memory CD4+ T cells, memory B cells, regulatory T cells, T follicular helper cells, Th17 cells, Th2 cells, activated CD8+ T cells, immature dendritic cells (DCs) and plasmacytoid DCs were significantly higher in the high-risk group, whereas the low-risk group had greater infiltration of activated CD8+ T cells and eosinophils (Figure 10A). In addition, the expression levels of all inhibitory immune receptors were higher in the high-risk group compared to that in the low-risk group (Figure 10B), indicating that the anti-tumor effect of high T cell infiltration was offset by a strong immunosuppressive tumor microenvironment due to overexpression of immune checkpoint proteins [18].
Cell assays: In vitro validation on DGUOK. HepG2 and MHCC-97H cells were treated with CCK8 and were performed to detect the cell viability. Hoechst 33258 fluorescent dye staining was used to show nuclear morphological changes and to assess apoptosis. Data in A are presented as means ± SD. **, p< 0.01. magnification: 200×.
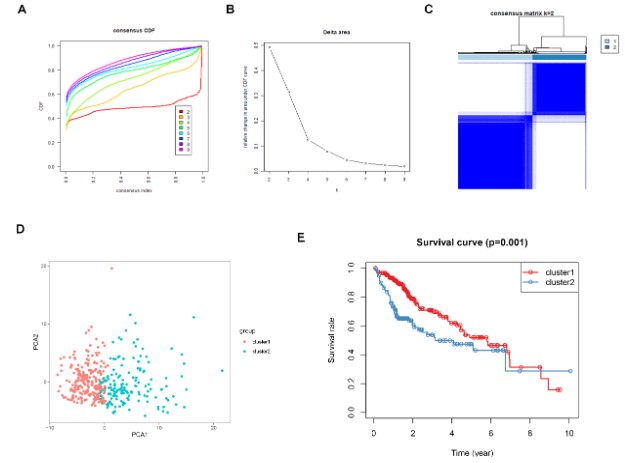
Figure 7: Consensus clustering for DDR genes in HCC patients from TCGA dataset. (A) The
cumulative distribution function (CDF) curve plot when k = 2 to k = 9 (B). The relative
change in area under CDF curve when k = 2 to k = 9. (C) Consensus heatmap when k =2. (D)
Principal components analysis for the DDR gene expression. (E) KM survival curve analysis
for the 2 subgroups.
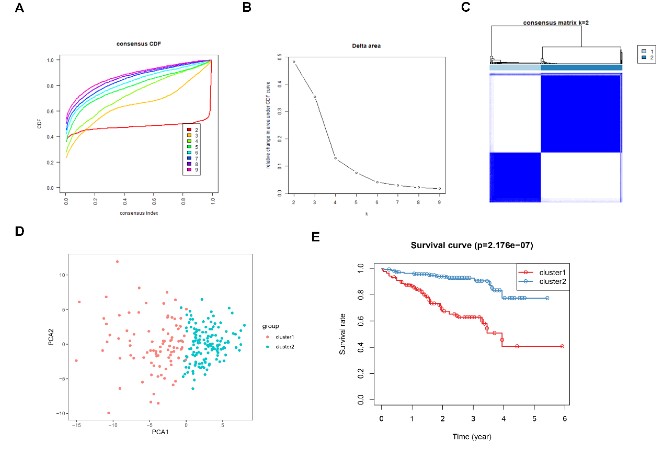
Figure 8: Consensus clustering for DDR genes in HCC patients from the ICGC cohort. (A) The
cumulative distribution function (CDF) curve plot when k = 2 to k = 9 (B). The relative change
in area under CDF curve when k = 2 to k = 9. (C) Consensus heatmap when k =2. (D) Principal
components analysis for the DDR gene expression. (E) KM survival curve analysis for the 2
subgroups.
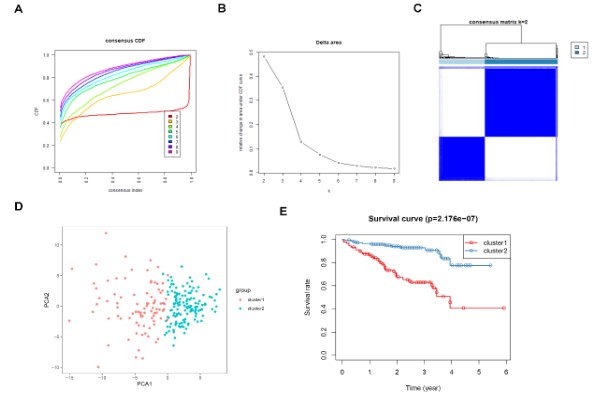
Figure 9: Heatmap analysis of the relationship between subgroup and clinical traits in
(A) TCGA and (B) ICGC cohorts.
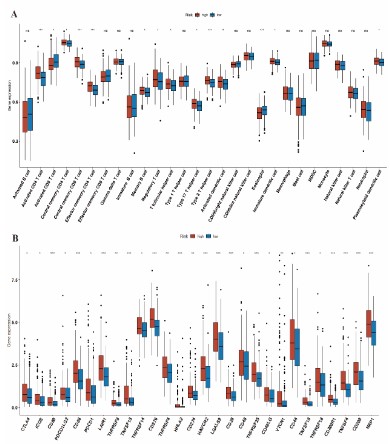
Figure 10: Immune infiltration. (A) The abundance of tumor infiltrating immune cells in high- and low-risk groups. (B) Boxplot showing the expression of immune-checkpoint genes in high- and low-risk groups.

Figure 11: In vitro validation on DGUOK. HepG2 and MHCC-97H cells were treated with siDGUOK for indicated time. A:
CCK8 was performed to detect the cell viability. B: Hoechst
33258 fluorescent dye staining was used to show nuclear
morphological changes and to assess apoptosis. Data in A are
presented as means ± SD. **, p< 0.01. magnification: 200×.
Discussion
HCC is a highly heterogeneous cancer with multiple risk factors such as hepatitis B/C, alcoholic liver disease, obesity etc. [19]. DNA damage and chromosomal aberrations are the initiating events of malignant transformation, and triggers a DNA damage response (DDR) in the affected cells. DNA lesions can be repaired through homologous recombination, mismatch repair, double strand break repair and other mechanisms. A dysfunctional DDR destroys genomic integrity and results in mutations, which eventually trigger carcinogenesis and promote tumor progression [20]. Studies show that DNA repair proteins such as sphingolipid signaling, hOGG1, XRCC1, TP53, PARP-1, MRE11-Rad50-NBS1 (MRN) complex and Ataxia Telangiectasia Mutant (ATM) kinase are frequently mutated in HCC [21]. In addition, the DDR pathway induced by ionizing radiation (IR) creates an immunosuppressive tumor microenvironment, which weakens the anti-tumor effect of radioimmunotherapy. DDR inhibitors can reverse the immunosuppressive state of HCC and inhibit tumor progression [22].
There is considerable evidence indicating that DDR genes directly influence tumorigenesis [23]. DDR genes are often aberrantly expressed in the tumor tissues or mucosa, and is closely associated with patient prognosis [24,25]. However, the prognostic value of individual genes is limited [26,27], and multigene signatures may be better suited for predicting HCC prognosis. However, no study so far has reported the prognostic value of DDR genes in HCC. We constructed a 11-DDR gene signature for predicting the prognosis of HCC patients using DDR gene expression data and clinical data from TCGA and ICGC databases. The risk score of the 11-gene signature demarcated the patients into the high- and low-risk groups, and the former was associated with worse survival outcomes. The risk model showed good predictive performance in both TCGA and ICGC datasets. In addition, the risk model was also identified as an independent prognostic factor of HCC. A nomogram constructed using the risk score and tumor stage clearly discriminated two prognostic groups, and may be useful in guiding preoperative management of HCC patients.
GSEA further revealed that the DDR gene signature was associated with cancer-related pathways including cell cycle, WNT signaling, mTOR signaling and apoptosis in the high-risk group, and are indicative of the potential mechanisms underlying HCC progression. The low-risk group on the other hand was enriched in metabolism-related pathways. Most of the genes in the DDR-based risk signature have an established role in tumorigenesis. CANT1 regulates pyrimidine metabolism in melanoma cells and is associated with tumor progression [28]. High CANT1 expression in prostate cancer cells is associated with better prognosis, and CANT1 silencing significantly suppressed cell proliferation and DNA synthesis [29]. CLP1, the first mammalian RNA kinase to be identified, plays an important role in motor neuron function [30]. Mitochondrial deoxyguanosine kinase (DGUOK) is the rate-limiting enzyme in the mitochondrial deoxynucleoside salvage pathway. Overexpression of DGUOK is associated with worse lung cancer prognosis, and its depletion suppressed lung adenocarcinoma growth, metastasis and CSC self-renewal [31]. GTF2B is a prognostic marker in neuroblastoma and colorectal cancer, whereas GTF2H1 is a p62 subunit of the multi-protein complex transcription factor IIH (TFIIH) and regulates nucleotide excision repair and transcription [32,33]. Some polymorphisms/haplotypes of GTF2H1 are associated with increased susceptibility to lung cancer [34]. Budding yeast orthologs of POLE4 enhance Polε processivity in vitro but are dispensable for viability in vivo and lead to accelerated tumorigenesis [35]. Moreover, the POLR2E rs3787016 polymor- phism may increase the risk of papillary thyroid carcinoma, prostate cancer, esophageal cancer, liver cancer and breast cancer [36-38]. However, the role of AAAS, NCBP2, POLA1 and POLR2D in HCC is unknown, and will have to be experimentally verified.
Immunotherapy has achieved encouraging results in various malignancies [39]. For instance, the “T+A” scheme is increasingly becoming the first-line option for advanced HCC [40]. Despite achieving good outcomes in multiple cancers, a significant percentage of the patients do not benefit from immunotherapy. Therefore, it is necessary to identify biomarkers that can predict the outcomes of immunotherapy, and screen for patients that can respond to immunotherapeutic regimens. Galon et al. [41] had proposed the concept of “cold” and “hot” tumors to evaluate their sensitivity to immunotherapies. In this study, we detected high infiltration of immunosuppressive cells and overexpression of immune checkpoint receptors in the high-risk group, which indicates that these patients are likely unresponsive to immunotherapy.
Conclusion
In conclusion, we developed an 11-DDR gene signature for predicting HCC prognosis. This prognostic signature can improve our understanding of the molecular mechanisms underlying HCC progression and guide clinical decision-making.
Declarations
Data Availability: The raw data could be obtained from the TCGA database. Additional Points: The “guarantor statement” does not apply to this manuscript.
Authors’ Contributions: YB contributed to data analysis, methodology, figures construction, and article writing. YW contributed to investigation and validation. JY H, YQ M, HL and ML contributed to methodology and validation. All authors contributed to the article and approved the submitted version.
Ethical Approval: Ethical Approval is not applicable for this article.
Statement of Human and Animal Rights: This article does not contain any studies with human or animal subjects.
Statement of Informed Consent: There are no human subjects in this article and informed consent is not applicable.
Funding: No funding.
Conflicts of Interest: The authors claimed that there were no conflicts of interest in this research.
References
- Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018; 68: 394-424.
- Davis GL, Dempster J, Meler JD, Orr DW, Walberg MW, et al. Hepatocellular carcinoma: management of an increasingly common problem. Proc (Bayl Univ Med Cent). 2008; 21: 266-280.
- Marengo A, Rosso C, Bugianesi E. Liver Cancer: Connections with Obesity, Fatty Liver, and Cirrhosis. Annu Rev Med. 2016; 67: 103-117.
- McGlynn KA, Petrick JL, London WT. Global epidemiology of hepatocellular carcinoma: an emphasis on demographic and regional variability. Clin Liver Dis. 2015; 19: 223-238.
- Lans H, Hoeijmakers JHJ, Vermeulen W, Marteijn JA. The DNA damage response to transcription stress. Nat Rev Mol Cell Biol. 2019; 20: 766-784.
- Nickoloff JA, Jones D, Lee SH, Williamson EA, Hromas R. Drugging the Cancers Addicted to DNA Repair. J Natl Cancer Inst. 2017; 109.
- Laurini E, Marson D, Fermeglia A, Aulic S, Fermeglia M, et al. Role of Rad51 and DNA repair in cancer: A molecular perspective. Pharmacol Ther. 2020; 208: 107492.
- Baretti M, Le DT. DNA mismatch repair in cancer. Pharmacol Ther. 2018; 189: 45-62.
- Mateo J, Boysen G, Barbieri CE, Bryant HE, Castro E, et al. DNA Repair in Prostate Cancer: Biology and Clinical Implications. Eur Urol. 2017; 71: 417-425.
- Wang X, Tan C, Ye M, Wang X, Weng W, et al. Development and validation of a DNA repair gene signature for prognosis prediction in Colon Cancer. J Cancer. 2020; 11: 5918-5928.
- Lin J, Shi J, Guo H, Yang X, Jiang Y, et al. Alterations in DNA Damage Repair Genes in Primary Liver Cancer. Clin Cancer Res. 2019; 25: 4701-4711.
- Iasonos A, Schrag D, Raj GV, Panageas KS. How to build and interpret a nomogram for cancer prognosis. J Clin Oncol. 2008; 26: 1364-1370.
- Vickers AJ, Elkin EB: Decision curve analysis: a novel method for evaluating prediction models. Med Decis Making. 2006; 26: 565-574.
- Wilkerson MD, Hayes DN. ConsensusClusterPlus: a class discovery tool with confidence assessments and item tracking. Bio-informatics. 2010; 26: 1572-1573.
- Hanzelmann S, Castelo R, Guinney J. GSVA: gene set variation analysis for microarray and RNA-seq data. BMC Bioinformatics. 2013; 14: 7.
- Charoentong P, Finotello F, Angelova M, Mayer C, Efremova M, et al. Pan-cancer Immunogenomic Analyses Reveal Genotype-Immunophenotype Relationships and Predictors of Response to Checkpoint Blockade. Cell Rep. 2017; 18: 248-262.
- Jia Q, Wu W, Wang Y, Alexander PB, Sun C, et al. Local mutational diversity drives intratumoral immune heterogeneity in non-small cell lung cancer. Nat Commun. 2018; 9: 5361.
- Matsushita H, Sato Y, Karasaki T, Nakagawa T, Kume H, et al. Neoantigen Load, Antigen Presentation Machinery, and Immune Signatures Determine Prognosis in Clear Cell Renal Cell Carcinoma. Cancer Immunol Res. 2016; 4: 463-471.
- Villanueva A. Hepatocellular Carcinoma. N Engl J Med. 2019; 380: 1450-1462.
- O’Connor MJ. Targeting the DNA Damage Response in Cancer. Mol Cell. 2015; 60: 547-560.
- Yang SF, Chang CW, Wei RJ, Shiue YL, Wang SN, et al. Involvement of DNA damage response pathways in hepatocellular carcinoma. Biomed Res Int. 2014; 2014: 153867.
- Sheng H, Huang Y, Xiao Y, Zhu Z, Shen M, et al. ATR inhibitor AZD6738 enhances the antitumor activity of radiotherapy and immune checkpoint inhibitors by potentiating the tumor immune microenvironment in hepatocellular carcinoma. J Immunother Cancer. 2020; 8.
- Ali R, Rakha EA, Madhusudan S, Bryant HE: DNA damage repair in breast cancer and its therapeutic implications. Pathology. 2017; 49: 156-165.
- Jinjia C, Xiaoyu W, Hui S, Wenhua L, Zhe Z, et al. The use of DNA repair genes as prognostic indicators of gastric cancer. J Cancer. 2019; 10: 4866-4875.
- Kun S, Duan Q, Liu G, Lu JM. Prognostic value of DNA repair genes based on stratification of glioblastomas. Oncotarget. 2017; 8: 58222-58230.
- Deng CX, Wang RH. Roles of BRCA1 in DNA damage repair: a link between development and cancer. Hum Mol Genet. 2003; 12: R113-123.
- Fang M, Xia F, Mahalingam M, Virbasius CM, Wajapeyee N, et al. MEN1 is a melanoma tumor suppressor that preserves genomic integrity by stimulating transcription of genes that promote homologous recombination-directed DNA repair. Mol Cell Biol. 2013; 33: 2635-2647.
- Edwards L, Gupta R, Filipp FV. Hypermutation of DPYD Deregulates Pyrimidine Metabolism and Promotes Malignant Progression. Mol Cancer Res. 2016; 14: 196-206.
- Gerhardt J, Steinbrech C, Buchi O, Behnke S, Bohnert A, et al. The androgen-regulated Calcium-Activated Nucleotidase 1 (CANT1) is commonly overexpressed in prostate cancer and is tumor-biologically relevant in vitro. Am J Pathol. 2011; 178: 1847-1860.
- Hanada T, Weitzer S, Mair B, Bernreuther C, Wainger BJ, et al. CLP1 links tRNA metabolism to progressive motor-neuron loss. Nature. 2013; 495: 474-480.
- Lin S, Huang C, Sun J, Bollt O, Wang X, et al. The mitochondrial deoxyguanosine kinase is required for cancer cell stemness in lung adenocarcinoma. EMBO Mol Med. 2019; 11: e10849.
- Di Pietro C, Ragusa M, Barbagallo D, Duro LR, Guglielmino MR, et al. Involvement of GTA protein NC2beta in neuroblastoma pathogenesis suggests that it physiologically participates in the regulation of cell proliferation. Mol Cancer. 2008; 7: 52.
- Ribeiro-Silva C, Aydin OZ, Mesquita-Ribeiro R, Slyskova J, Helfricht A, et al. DNA damage sensitivity of SWI/SNF-deficient cells depends on TFIIH subunit p62/GTF2H1. Nat Commun. 2018; 9: 4067.
- Wu W, Liu H, Lei R, Chen D, Zhang S, et al. Genetic variants in GTF2H1 and risk of lung cancer: a case-control analysis in a Chinese population. Lung Cancer. 2009; 63: 180-186.
- Bellelli R, Borel V, Logan C, Svendsen J, Cox DE, et al. Polepsilon Instability Drives Replication Stress, Abnormal Development, and Tumorigenesis. Mol Cell. 2018; 70: 707-721 e707.
- Chen B, Li J, Yi C, Jiao Y, Gu X, et al. Long non-coding RNA POLR2E rs3787016 is associated with the risk of papillary thyroid carcinoma in Chinese population. Pathol Res Pract. 2018; 214: 1040-1044.
- Chen B, Jiao Y, Yaolong F, Li T, Liu Y, et al. The POLR2E rs3787016 polymorphism is strongly associated with the risk of female breast and cervical cancer. Pathol Res Pract. 2019; 215: 1061-1065.
- Chen B, Wang S, Ma G, Han J, Zhang J, et al. The association of POLR2E rs3787016 polymorphism and cancer risk: a Chinese case-control study and meta-analysis. Biosci Rep. 2018; 38.
- Yang Y. Cancer immunotherapy: harnessing the immune system to battle cancer. J Clin Invest. 2015; 125: 3335-3337.
- Finn RS, Qin S, Ikeda M, Galle PR, Ducreux M, et al. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N Engl J Med. 2020; 382: 1894-1905.
- Galon J, Bruni D. Approaches to treat immune hot, altered and cold tumours with combination immunotherapies. Nat Rev Drug Discov. 2019; 18: 197-218.
